- ________________________________________________________________________________________
- Last modified 03/03/08
- Copyright © by Nila Gaede 2008
| The conveyor belt theory of extinction |
| Adapted for the Internet from: Why God Doesn't Exist |
- 1.0 You cannot explain away periodic extinction patterns with comets
Most analysts seem to agree that our planet has suffered the extinction of as much as 99% of the animal species that ever
lived. The most prominent of these extinctions are the mass extinctions, a very short geological period in which a broad
category of species disappear. The mighty dinosaurs of the Late Cretaceous and the ferocious synapsids of the End
Permian readily come to mind. What most analysts seem to overlook is that these extinctions claimed a very broad
category of plants as well. Just for starters, here are some suspicious examples:
- • Cretaceous. T-Rex fed on Triceratops which likely fed on cycads and cycadeoids.
- (p. 171) [1] The entire system vanishes towards the end of the Cretaceous.
The staple diet of Mosasaurs consists of mollusks, primarily ammonites, [2] [3]
which mostly feed on particular types of plankton, likely the ones that
disappeared at the K-T boundary: foraminifera, diatoms, and calcareous
nannoplankton. [4] [5]
- • Jurassic. Allosaurus indulged in long-necked Sauropods which subsisted on
- the towering conifers -- perhaps ancient araucaria -- that are no longer around.
- • Triassic: The archosaur Prestosuchus hunted the rinchosaur Hyperodapedon
- which probably grazed on Dicrodium (seed fern). This particular system
disappears around the Carnian subdivision of the Triassic. There is no evidence
that the dinosaurs displaced the archosaurs because of superior adaptability or
might as argued by many. [6]
- • Late Permian: Gorgonops fed on Pareiasaurus on the South African Karoo while
- Inostrancevia fed on Scutosaurus in Siberia. The Permian interval is characterized
by a changing climate from a humid to dry and from lower to higher oxygen
atmosphere. This may have helped the incoming gymnosperm (cycads, conifers)
displace the ruling glossopteris, but probably the glossopterids had already been
around for too long and were undergoing the universal process of aging. If
Pareiasaurus was the most abundant herbivore and Glossopteris the most
abundant plant in Gondwana, and Gorgonops and Pareiasaurus probably grew in
a symbiotic relation, it's very likely that the entire system disappeared when these
plants vanished at the end of the Permian.
- • Early Permian: Edaphosaurus and calamites lived throughout the same period,
- from the Carboniferous to the end of the Early Permian. The sail on the back of this
species closely resembles a calamite, which both Edaphosaurus and his cousin
{kind=link}
Dimetrodon used as camouflage against each other. And if not calamites, then
{kind=link}
perhaps either Lepidodendron and Sigillaria, primitive lycopods that disappeared
around the same time.
- likely were feeling density pressures as the incoming seed ferns were gradually
displacing the more archaic fern allies.
Fig. 1 Synopsis of the history of terrestrial dynasties [You will find a more accurate history including number of species per major plant category here.] |
Referring to the Cretaceous, Russell comments that:
- “ As shown by the work of Sheehan and Fastovsky, the extinction seems to be
linked to animals that depended on foliage for food, and the carnivores that in
turn depended on the foliavores for food.” [7]
I could also have subdivided the Phanerozoic Eon (the interval in which ‘animals’ have lived on Earth) into the same three
eras and renamed them: the Age of Trilobites (Paleozoic), the Age of Ammonites (Mesozoic), and the Age of Krill (Cenozoic)
(Fig. 2). In this particular scheme, I would be dividing the Phanerozoic in terms of the most important intermediary between
plankton and medium to large oceanic predators. Once the plankton are gone, the entire system is gone.
3.0 Why terrestrial and aquatic life dies simultaneously
More significantly, the conveyor belt view of life explains why land and sea creatures massively die at about the same time.
Terrestrial and oceanic plants and animals subjected to the same temperatures throughout their histories enjoy a symbiotic
relationship through the atmosphere. They gradually tailor the environment to suit their needs, but this ends up killing them.
To get a feel for what I'm talking about, think of modern Man. Let's assume we continue to pollute the atmosphere without
any restraints. More people, more pollutants. Let's further assume that we expand demographically for the next few thousand
years. At some point, many of the atmosphere parameters will reach critical levels. The ozone might break down, or the
greenhouse effect enhanced due to excessive CO2 in the atmosphere. In other words, by simply living our routine lives, we
would be digging our own graves.
Visualize the world in the middle of the Permian. There is high oxygen contact in the atmosphere, perhaps as high as 30 or
35 %. In comparison, today there is only 18%. By the beginning of the Triassic, the oxygen level falls to lowest ever level, to
about 13%. Throughout the rest of the Mesozoic, the oxygen increases gradually to about 22 %, almost twice as much as
there was at the beginning. Then just before the Cretaceous is about to expire, the oxygen level dips dramatically again to
17%. If plants are the main contributors to atmospheric oxygen, it means that plants increased in volume throughout most
of the Mesozoic and that animals were never able to keep up. Then at the end of the Cretaceous, there is a sudden reversal.
The volume of plants goes down and the volume of animals goes up.
Add now the nitrogen cycle. This is perhaps the most critical biological pump that drives life on Earth. The type of sea and
terrestrial life that develops is a result not only of temperature, but of chemical resources. If there is more nitrogen in the
atmosphere, a nitrogen-fixing plant undergoes a golden age. When nitrogen depletes, we can expect the populations of sea
and terrestrial plants to run into trouble. By shear coincidence cycads and cycadeoids as well as phytoplankton (tiny sea
plants) known as diatoms and calcareous nannoplankton are nitrogen fixing plants. All of these – both terrestrial and
aquatic – flourished from the Jurassic to the Cretaceous. Certainly, a reduction in atmospheric nitrogen would have
adversely affected them. Or vice versa. The fact of atmospheric nitrogen depletion indicates that the nitro-plants were
disappearing. On land, the record shows that a great diversity of animals appear in the Cenomanian (Late Cretaceous,
95 mya) just when the cycads and cycadeoids are turning the corner. This is a period in which nitrogen is declining. There
is a time lag between the expansion of plants and the expansion of animals. As the gymnosperms are receding, the
dinosaurs are incongruously multiplying in species and in numbers. The Golden Age of Dinosaurs is happening when the
Age of Cycads has run its course. Predictably, the dinosaur families reach a carrying capacity plateau. Soon after, they
begin to vanish by attrition. Radiation stops because there is no new niche to carve along this now thinning conveyor
belt. The remaining species meanwhile specialize, become mightier, and each one continues towards the end of its
branch. By the end of the Cretaceous, the numbers of dino species have dwindled to a handful and the cycadeoids dip
out of sight in a sea of angiosperms. We must conclude that the evolution of diatoms, foraminifera, calcareous
nannoplankton, and cycadeoids is inextricably linked to nitrogen and formed the basis of the food chains that disappeared
with them.
One effect of these trends is a change in temperature. If the beginning of the Mesozoic (the Triassic) was very hot, the end
of the Mesozoic (the Cretaceous) is comparatively cold. The temperature and the variable oxygen - CO2 ratios don't kill the
plants and animals as many nitwits propose. What they do is place physical limits on where terrestrial and oceanic plants
can migrate. You will not see cycads naturally migrating to the poles! The role of temperature is to create islands, to form
an invisible fence beyond which certain plants cannot migrate. Once the cycads and cycadeoids (on land) and
coccolicophores and diatoms (in the sea) are constrained by temperature, they reach their maximum extent and their
population pyramid overturns. These plants have now been around for millions of years. They have fought disease and
developed antidotes against herbivores along the way. In the end, they just postpone the day of reckoning. The
herbivores also develop ways to counteract and overcome the poisons and thorns and other defenses. The population of
plants expands to a limit and now a decline in fertility takes place. It takes place at the wrong time because by now the
herbivores have also grown in size and in numbers although fewer species are left. The plants are declining in numbers
just when the mass of herbivores is exploding. A critical point is reached where plant reproduction cannot keep pace
with destruction. Herbivores are now proactively digging their graves. Enter the carnivores. They can only expand
subsequent to the expansion of the herbivores. There is a time lag between the plants and the herbivores and another
one between the herbivores and the carnivores. The old species of plants are vanishing when the old species of
herbivores are increasing in mass (numbers and size), soon to be followed by an increase in the mass of carnivores.
The entire system is doomed. We should see in this movie two critical points. One where the herbivores run into trouble
with the declining plants and another one shortly after where the carnivores run into trouble with the now declining
herbivores. These free-falls should be exponential.
Therefore, without going into detail, from a strictly conceptual point of view, we can expect the biological cycles and the
temperatures on Earth to gradually change until they reach critical proportions for both oceanic and terrestrial life that
mediate the cycles. In the long run, the plants are working against their own interests. The plants destroy the habitat that
provides life to them just like Man today destroys the habitat which he requires to remain alive. When the basic staple of
the large animals was removed from them both on land and in the sea, the entire system collapsed. The new breeds of
plants and animals help by requiring other needs.
In the Carboniferous, the mass of plants was great and the mass of animals was comparatively small. The result is that
the carboniferous had a high oxygen/low CO2 atmosphere. In the long run, this certainly worked against the non-vasculars
and early vasculars -- accustomed to a low oxygen atmosphere -- and helped the incoming ferns and gymnosperms. [30]
Likewise, by the mid-Cretaceous, the level of CO2 had gone up and the oxygen had dropped, sealing the fate of the
gymnosperm to the benefit of the flowering plants (angiosperm).
The first question that biochemical pumps answer is the crucial issue of 'globalization.' If a mass extinction affected similar
plants and animals in different parts of the world, there is no better, foolproof mechanism than a biological pump.
- " Among the many black-shale horizons identified on land and in the oceans, the
Selli Event (OAE1a) and Bonarelli Event (OAE2) have proven to be of global
distribution. Both are recorded, for example, on submarine plateaus in the Pacific
Ocean. In 2002, the black shale of the Selli Event was drilled on Shatsky Rise
during ODP Leg 198. This extremely carbon-rich horizon (~35% TOC) contains
biomarkers for cyanobacteria that could have utilized atmospheric elemental
dinitrogen if nitrate levels in the photic zone became vanishingly low because of
utilization by plankton (Brassell). This discovery points to upwelling and high
productivity as a major forcing of OAE1a. " [31]
From the middle of the Cretaceous onwards, the Carbon content in the atmosphere increased at the expense of nitrogen.
Coincidentally, the aquatic and terrestrial primary production that disappears at the K-T boundary (diatoms, calcareous
nannoplankton, foraminifera, cycads, cycadeoids) are heavily dependent on the nitrogen-cycle. We should expect these
living entities to run into trouble as the environment they thrive in changes. If during the same period, the animals that
depend on these sources increase in numbers and in size (meaning that the volume they consume increases), at some
point the system has to reach critical proportions and subsequently collapse exponentially. The ecological pyramid turns
abruptly because we have the many chasing after the few.

| God's workshop |
- 4.0 Conclusions
Plants belong to the world of the living and, therefore, their demography should be subject to the sigmoidal ‘S’ curve too.
This means that they must also experience a long period of arithmetic growth followed by a comparatively shorter period
of exponential growth followed by rapid decline. Animal species fated to live during the declining phase of their favorite
food are unfortunately at the wrong place at the wrong time. New classes of plants are besieging and choking off the
declining empire. As plant species of the old order die out, medium and large animal diversity decreases through a
process of attrition. Faunal species vanish along the way when their demographic pyramid overturns. New ones do not
radiate because the traditional environment is shrinking. Instead, the old ones evolve into ever thinning branches. This
process results in fewer, more formidable species which remain faithful to the ancient regime. Only the best make it
through the bottleneck. The successful species have by now had time to conquer diseases and attain their natural life
spans, grow in size and in numbers, and develop voracious appetites. The now streamlined herbivores are unwittingly
aiding the incoming plants in their conquest of the land. Here and there a few species manage to carve a niche in the
incoming families of plants which are largely the domain of a new class of animals too tiny to overthrow the prevailing
order. Hence, it is not surprising that large animals are more prone to extinction despite that they have succeeded in
outmaneuvering smaller ones. In this context, extinction is not really related to size. It has to do with the species’ location
on the dynastic chronology. If we superimpose on these graphs the grand evolution of plants, we can more or less
correlate the fall in animal diversity to the disappearance of ancient classes of plants.
Granted, I am oversimplifying a complex subject enormously. Certain types of gymnosperm known as seed ferns were
already numerous in the Carboniferous [8] and non-vasculars can probably be traced to the first plants on land. Likewise,
the Therapsids dominated during the Late Permian, but they evolved from Pelycosaurs. And the Sauropsids took over
from the declining Archosaurs during the Triassic. The point of Fig. 1 is to focus on the forest without being distracted
by the trees. We must concede that there was an evolution of plants from the most primitive to the ones we see today.
The early non-vasculars had no seeds, leaves, or internal system to channel water and nutrients. The angiosperms do.
Therefore, not only is a non-vascular a radically different kind of plant than an angiosperm, but without a doubt also a
more primitive one. Before Mother Nature could create the angiosperm, she had to toy around with non-vasculars,
fern allies and ferns (spore reproduction), and gymnosperms (cone reproduction). Likewise, we can be sure that a bear
is a much more evolved animal than was either Cacops or Dimetrodon who lived hundreds of millions of years ago. The
primitive species of animals that appeared on land had to develop the basics: hearing, walking, making sounds, birthing,
etc. A certain, broad family of plants dominated the landscape for lengthy periods of time and served as food for the
majority of herbivores which in turn served as food for carnivores. If at some chronological point these plants came to an
end, the species of animals that depended on them must have come to an end also. Of course, a few late bloomers of the
old guard may carve out a niche within the new classes of plants and animals, and this may explain some of the stragglers
we find on the other side of the divide now and then (e.g., a few species of trilobites crossing the Permian, [9] the
cynodonts crossing the Triassic).
The purpose of this simplification is to get the basics right, because the so called 'experts' seem to ignore them completely
during their dissertations. If we were to stand at the start of the Cambrian, this 'conveyor-belt' mechanism enables us to
predict that broad categories of plants together with the animals that depend on them will periodically disappear. As an
analogy, visualize a diverse fauna sitting on a conveyor belt that expands as we roll it forward (Fig. 3). More varieties of a
grand family of plants develop as the belt widens. The numbers of species and quantities of animals per species increase
in proportion to this expansion. At some point the plants have run their course and both the varieties and numbers of
these families of plants begin to decline. We can also expect the families of animals tied to the old order to crash when
these plants come to an end. We should expect to see at some point ever more numerous herds of ever larger specimens
of ever fewer species subsisting on ever scarcer vegetation.
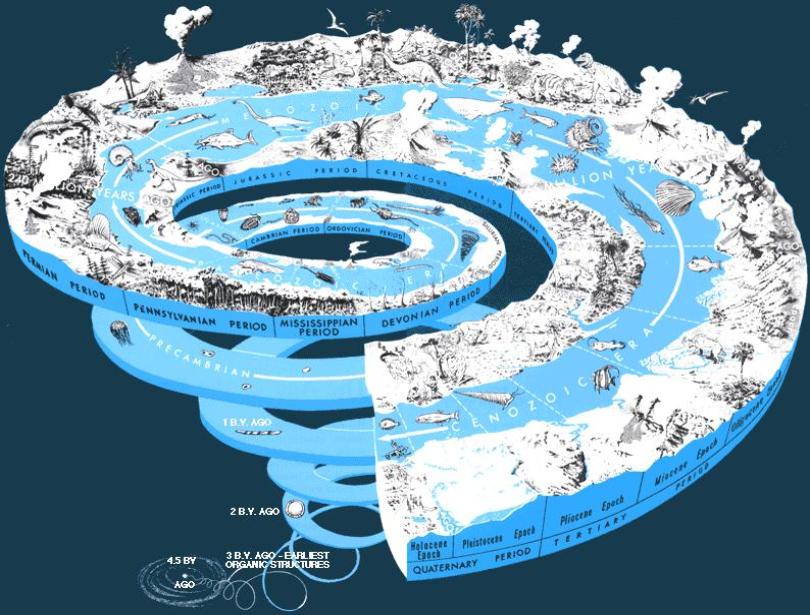
Fig. 2 The Phanerozoic |
| Each of the three eras of the Phanerozoic can be characterized in terms of the dominant trophic level which served as a link between plankton and larger animals. |
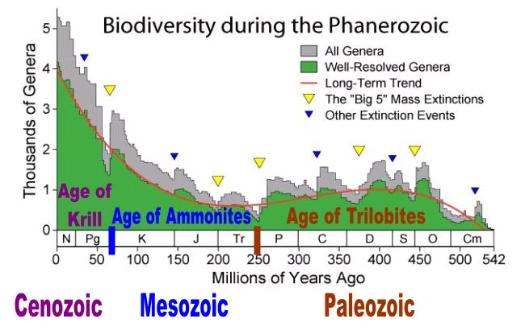
Catastrophic theories (especially those that rely on extraterrestrial agents) do not account for the general patterns depicted
in Figs. 1, 2, and 3. Left to Raup and Sipkoski, we would never get to see a 'natural' mass extinction because their angry gods
always end up selectively destroying archaic life with one of their asteroids or comets. Or we would never be able to explain
what some lame-brained paleontologists still question: Cope's Law. Why is it that mass extinctions always and without
exception take out a bunch of monsters?
- " Parapuzosia seppenradensis is the largest known species of ammonite. It lived
during the Late Cretaceous period. A specimen found in Germany in 1895 measures
1.95 m in diameter" [10]
" The partial shell of one giant Cameroceras yielded a total length estimated at the
time at nearly 30 feet." [11]
" Dunkleosteus... lived in the late Devonian period... The largest of the genus grew
to around 8 to 10 m (27 to 33 feet) long, and were probably the top predators of
their time." [12]
- " the huge green-stemmed Lepidodendrale lycopods (Lepidodendron, Sigillaria, etc,
35 meters tall), the giant sphenopsid Calamites (20 meters in height), and the
strap-leaved mangrove-rooted Cordaitales (Cordaites, up to 45 meters) are all
abundant" [13]
" The dragonfly-like Meganeura, an aerial predator, had a wingspan of 60 to 75 cm.
The inoffensive stocky-bodied and armoured millipede-like Arthropleura was 1.8
meters long, and the semi-terrestrial Hibbertopterid eurypterids were perhaps as
large" [14]
" Dimetrodon was the dominant predator in its environment for some twenty-five
million years, during which time it evolved through about a dozen species,
become steadily larger as time progressed (an example of "Cope's Law"). [15]
" Some of the segments of trunk represent giant trees that are estimated to have been
over 50 meters tall when they were alive." [16]
" Before the [K-T] mass extinction, most of the foraminifera species were comparatively
large... roughly 200 to 350 microns large, or a fifth to a third of a millimeter long." [17]
" delayed maturation and growth to large sizes are only advantageous under stable
environmental conditions where food resources are limited… These traits are
sometimes associated with increased embryon size and suppression of sexual
reproduction… Specialization for these traits…often dooms the spieces or lineage
to extinction" [18]
- " as long as the large carnivore ecospace was filled, the radiation of new taxa into that
ecospace was limited, only occurring after the extinction of the incumbents. The
apparently inevitable decline of incumbent taxa may reflect the tendency for clades
of large carnivorous mammals to produce more specialized species as they mature,
leading to increased vulnerability to extinction" [19]
- I could go on and on with the examples of gigantic species animals that did not disappear in their evolutionary infancy
-- Triassic archosaurs, Jurassic Camarasaurus, Cretaceous T-Rex, etc. Why doesn't a major mass extinction for once take
out the little guys?
- " Fossil taxa in the immediate aftermath of mass extinction events are much smaller
than in the pre-extinction fauna. This 'Lilliput effect' is widespread and has been
documented in the aftermath of many extinction events and affects many different
groups of organisms." [20]
Why doesn't the stupid comet strike when a few species are developing right after the last mass extinction took out the big
guys?
What is even harder to explain with extraterrestrial agents is that mass extinctions always and without exception target the
ancient species of plants and animals. These smart comets have a magical ability to distinguish between the archaic and
the upwardly mobile. It is this chronological selectivity that is suspicious and which is difficult to justify with catastrophic
theories.
2.0 Evidence
We see evidence for the conveyor-belt theory throughout the fossil record. Bakker synthesizes the chronological distribution
of species in terms of biomass for each geologic epoch. [21] [22] If his graphs more or less depict what actually occurred, we
see a repeating, grand pattern in the history of life. For each major epoch, there is a long initial period with relatively few
species followed by a shorter period containing many species (Fig. 3). The last phase shows a geologically rapid shrinkage
by attrition in which Cope’s Law is furtively at work. Few species consisting of large animals and relatively large populations
remain. Shortly after, they vanish as well. Bakker provides at least two examples that support my argument:
- 1. The long-necked dinosaurs increased in size throughout the Jurassic in response
to the tall conifers that were populating the land. (pp. 185-187) [23] The particular
conifers of those days perhaps disappeared, but if not, they moved to higher
- latitudes (another macro tendency of plants that we see in the record). The next
period, the Cretaceous, is characterized by low grazers probably munching on
cycadeoids (Fig. 4). It is tentative to conclude that the extinction of the long-necks
has something to do with the extinction of the tall plants they relied upon.
- 2. The Judith River fauna in Northwestern USA during the last stages of the
- Cretaceous consisted of a healthy mix of duckbills and horned dinosaurs. The
next layer, the Scollard, is more uneven, with the duckbill Saurolophus consti-
tuting 75% of big specimen. In the final Edmonton-Hell Creek level, it is the horned
Triceratops that makes up to 80% of large dinos (pp. 435-439). Shortly after, the
Triceratopses are gone. Some researchers suggest that Triceratops was built to
eat cycads (e.g., Ostrom and Bakker, p. 171). I will add to this flora the cycad-like
plants -- the cycadeoids -- which abounded in the region and coincidentally
disappeared with Triceratops. We know for sure that South Dakota had great
forests of cycadeoids because one of these was fossilized and survived until our
days. Apparently, Triceratops went left instead of right, drifted to Montana, and
missed this bed.
F
- The cycadeoids were a small percentage of the vegetation at the end of the Cretaceous. [24] [25] 90 % of the plants were
angiosperm. I infer from this data that dinosaur herbivores did not eat magnolias. Flowering plants were weeds as far as they
were concerned. The dinos had not evolved a relation with this flora.
Some paleontologists argue that herbivores will change their diets when food goes scarce. For instance, Stromberg argues
that Late Cretaceous sauropods ate grasses:
" Silicified plant tissues (phytoliths) preserved in Late Cretaceous coprolites from India
show that at least five taxa from extant grass (Poaceae) subclades were present on
the Indian subcontinent during the latest Cretaceous... Other phytoliths extracted from
the coprolites (from dicotyledons, conifers, and palms) suggest that the suspected
dung producers (titanosaur sauropods) fed indiscriminately on a wide range of
plants. [26] [27]
- It is difficult to accept that this is the case. First, it is unlikely that an animal that is already present changes its diet from cycads
and conifers to grasses. There are plants and there are plants. Certainly, you don't switch to weeds and pine cones when you
can't find lettuce and apples. Assuming that cycads and conifers become scarce, the dinosaurs would have adjusted their
numbers to the available food and not spit our children irresponsibly, hoping that they will adjust their diet to a new type of
plants. Another potential problem is that most paleontologists seem to agree that grasses did not appear until the Cenozoic.
The most difficult argument to overcome, however, is that processing of grasses requires that the animal have a well
developed rumen and the bacteria to come with it that help digest the grasses. Again, it is difficult to believe that the last
sauropods, who had a long tradition of eating conifers and cycads suddenly developed the ability to process grasses.
Russell argues that the shear size of sauropods would have made the digestion process impossible:
- " Acetic acid (the acid found in vinegar) is the primary end-product of rumen
fermentation, and the ruminants use this acid for energy. If the rumen retention
time gets too long, a bacterium called Methanosarcina is able to convert acetic
acid to methane and carbon dioxide. Since these gases are belched away, the
animal would loose most of its energy. Nature has prevented the problem of
acetate conversion to gas by making sure that retention time is never too long
and is short enough to wash Methanosarcina out of the rumen. The problem with
dinosaurs having a traditional rumen is their very large body size. The largest
plant-eating dinosaurs weighed more than 25 tons. Based on the observation
that feed retention time in the rumen and body size appears to be a linear function,
the retention time of a dinosaur rumen could be greater than 1250 hours, a time
clearly not compatible with efficient acetate utilization." [28]
- Sullivan makes a recount of dinosaur species that lived throughout the Mesozoic:
" For dinosaur species: late Campanian: 103 maximum, 77 minimum; early Maastrichtian:
75 maximum, 68 minimum; late Maastrichtian: 49 maximum, 46 minimum. For dinosaur
genera: late Campanian: 92 maximum, 69 minimum; early Maastrichtian: 67 maximum,
60 minimum; late Maastrichtian: 46 maximum, 43 minimum. In either case, there is a
noticeable decline of taxa in both the genera and species from the late Campanian to
late Maastrichtian."
[Campanian (83 to 71 mya) and Maastrichtian (71 to 65 mya) are the last two stages of the
Cretaceous.]
- If his numbers have any validity, there was already a general mass extinction underway. More species of dinosaurs were
disappearing in the last 10 million years of the Cretaceous than were coming aboard. Meanwhile, the new breeds of tiny
mammals (multituberculata [22]) crawling at their feet, the crocs, and the turtles are unaffected by all the commotion. The
fast-swimming, well-adapted mosasaurs, the humongous, flying Quetzalcoatlus, and the archaic Bennettitales die out, but
the modern birds and magnolias survive the alleged asteroid collision, the ensuing impact winter, and expand unrestrainedly
into the Cenozoic. [29] We must conclude that, more than large, the asteroid must have been magical if it could discern
between the ancient species of plants and animals and the new. People are really gullible and ignorant if they believe that
the Cretaceous extinction is going to be resolved with an extraterrestrial impact! We should put theories that come from
contemporary mathematical physicists where they belong: in the toilet!
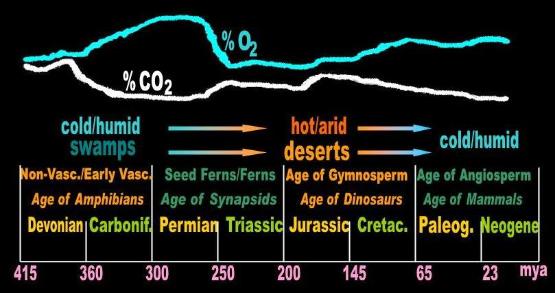
Fig 4 Long necks and conifers are followed by T-Rex, low grazers, and cycad/cycadoids. All disappear from North America at the end of the Cretaceous. |
| Tall conifers and long necks |
| T-Rex, Triceratops, and cycadeoids |
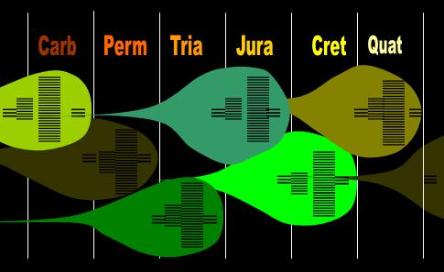
Fig. 3 The Conveyor Belt Theory of Mass Extinctions |
Module main page: Mass extinctions: How the mighty T-Rex really vanished
Pages in this module:
- 1. A mass extinction results when the ecological pyramid overturns
2. This page: The conveyor belt theory of extinction
3. The ecological pyramid overturns: a case study
4. A brief history of homosexuality
| As new species of plants (balloons) develop, new species of animals (lines) carve niches in them. The subsequent demographic and diversity expansion of this plant carries with it a greater population and diversity of animals. It can do so only at the expense of the established order. The previous rulers of the plant kingdom have reached their peak some time ago and are now undergoing a pyramid inversion. Incongruously, the animal species that depend on them and which rule the planet are experiencing demographic and diversity expansion |
| just when the plants on which they depend are suffering intense competition from new arrivals. Towards the end of an era, only few species of highly specialized, formidable animals develop consistent with Cope’s Law. These latebloomers and their ancestors ironically have gradually conditioned the landscape for the new breeds that will inherit the planet when the old guard dies off. |
